Mesothelial-Mesenchymal Transition (MMT) Mechanism
| ✅ Paper Type: Free Essay | ✅ Subject: Biology |
| ✅ Wordcount: 3716 words | ✅ Published: 08 Jun 2018 |
Mesothelial cells play a vital role in maintaining the structure of peritoneal membrane and peritoneal homeostasis. During long term peritoneal dialysis, prolonged exposure of peritoneum membrane to peritoneal dialysis solution causes peritoneal fibrosis. Bioincompatible dialysis fluid causes mesothelial cells to undergo uncontrolled mesothelial-to-mesenchymal transition triggered by Transforming growth factor-β1, which leads morphological and functional disruption of membrane. This literature review is about mechanism of mesothelial -mesenchymal transition (MMT), myofibroblasts and its origin hypothesis, clock genes and their cross relation with organ fibrosis.
1.1 Peritoneum anatomy
Peritoneum is defined as gliding membrane, composed of mesothelial cells maintained by basement membrane and thin layer of connective tissue. It consists of two distinct layers of mesothelium, known as parietal peritoneum and visceral peritoneum. Parietal peritoneum lines the peritoneal cavity and comes from somatic mesoderm of the embryo. Visceral covers the external surface of most abdominal organs and derived from splanchnic mesoderm of the embryo. Peritoneal cavity is potential space between both layers and is comprised of small amount of lubricating fluid, also known as serosal fluid which prevents friction between closely packed organs and allows free movement of them. [1,2] The primary function of peritoneal membrane is to provide support to abdominal organs. It also acts as passage to their blood vessels, lymph vessels and nerves.
Peritoneal cavity is rich in number of cells like mesothelial cells, fibroblasts, endothelial cells, adipocytes and immune cells like macrophages, T cells, B cells, mast cells. Along with cells, they contain cytokines and growth factors, like interleukin-1 (IL-1), Transforming growth factor-β1 (TGF-β1), Fibroblast growth factor (FGF), and Hepatocyte growth factor (HGF). All through the development process of peritoneal fibrosis, peritoneal cells will experience changes and some of the pro inflammatory factors which act as inducers to induce fibrosis and inflammatory processes. In this designed project, we are going to focus on mesothelial cells along with Transforming growth factor-β (TGF-β1), which is considered as main mediator of organ fibrosis development.
1.2 Peritoneal Fibrosis caused by peritoneal dialysis
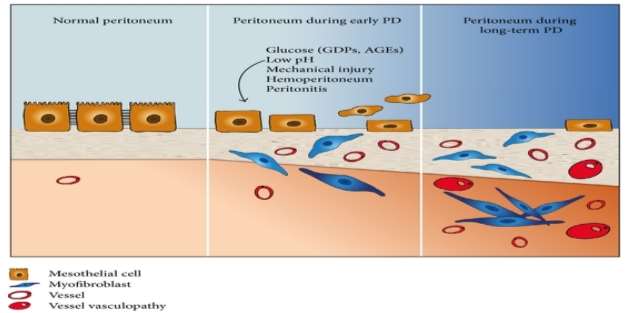
Peritoneal Dialysis, an alternative to haemodialysis is being used about more than three decades ago to treat end stage renal disease. During this treatment, PD solution is introduced into the abdomen with the help of tube also known as catheter. This dialysis therapy uses peritoneum membrane as semi-permeable membrane and principle of diffusion, to diffuse waste from underlying blood vessel into the solution.

PD solution is major drawback of this treatment which contributes to development of peritoneal fibrosis in patients due to its bioincompatible nature. Long-time exposure to hypertonic environment of PD solution with high glucose concentration, acidic pH, presence of Glucose Degradation Products (GDPs) and Advanced Glycation end-products(AGEs) causing functional and structural alterations in peritoneum membrane. [43]
Other factor which triggers fibrosis process in peritoneum membrane during dialysis is peritonitis. Peritonitis is inflammation of peritoneum which is caused by infection that can occur either by using unsterile catheter tube, handling which lead to introduction of micro-organism into abdominal cavity. Interesting research was done by Troidle and team, where they reported that inflammation caused by Gram-positive bacteria in patient peritoneum are better managed than the one caused by gram-negative. [81]
Numerous clinical studies carried out to study changes that occur in peritoneum of patients undergoes peritoneal dialysis. Till date, largest peritoneal biopsies study was carried out by William and colleague on total of 212 subjects, out of which 9 were normal individuals, 48 undergoing hemodialysis, 25 uremic and 130 subject undergoing PD. [44] There submesothelial thickness was measured, also were checked for vasculopathy and general morphological features. For normal individuals, submesothelial thickness was 50 ïm, patients undergoing hemodialysis and uremic patients was 140 ïm and 150 ïm respectively and for PD patients, thickness was 270ïm. Similar thickness of uremic and haemodialysis patients indicates that uremia can cause change in the peritoneal membrane. Value of submesothelial thickness increased to 700 ïm in long term PD patients, which directs towards fibrotic response of peritoneal to PD. They were also able to identify vascular changes including increasing hyalinization leading to vasculopathy. These findings were confirmed by Plum et al and were able to identify link between increasing thickness of submesothelial layer and increasing transport rate of solutes. [45]

1.3 Mesothelial Cells – Structural and Functional Properties
Even though being mesoderm origin, mesothelial cells possess various characteristics of epithelial cells. Normally, mesothelial cells are squamous, and flattened in morphology. They are rich in cell organelles, microvilli and cilia are present on the surface. While cilia aim to regulate surfactant secretion, microvilli functions to entrap water and serous exudates in order to guard cell surface from frictional injuries. [5]
Glycocalyx created by glycosaminoglycan, proteoglycans, and phospholipids surround mesothelial cells and have dual function. They form non-sticky slippery surface which supports intracoelomic movement and also provide protection against scratch.
Mesothelial cells are specialized cells which plays key role in peritoneal homeostasis and maintains the integrity of the peritoneal membrane. The critical roles of mesothelial cells in peritoneum are to secrete and synthesize growth factors, cytokines and chemokines in peritoneal cavity by transport of fluid and solute across the membrane. [6]
Mesothelial cells are considered as cells that are first in line in body’s defense system and they are able to sense and responds to signals within their microenvironment. They actively participate in deposition and clearance of local fibrin in the serosa membrane cavities. After mechanical injuries and infections, their fibrinolytic activity is important to prevent and remove any fibrin deposits that may forms. [7] Peritoneal membrane does act as selective permeability barrier to the channel of plasma proteins due to existence of anionic sites on mesothelial cells which in result provide net negative charge to the peritoneum membrane.
In normal conditions, mesothelial cells are identified on its ability to express cell markers like occludin, claudin-1, ZO-1, Wilms Tumor-1, and E-cadherin. When mesothelial cells undergo Mesothelial-to-mesenchymal transition which we will be explained in detail later; they either unable to express them or their level expression decreases. Therefore, it’s important to know about all mesothelial cell markers for easy identification within pool of cells.
1.3.1 Cell markers
Cell markers are defined as molecules with distinctive genetic and biochemical characteristic. These cell markers are present in cells and can be either proteins, peptides or carbohydrates and mainly present in the cells. They are used to distinguish between different cell types. Some of cell markers are specific to cell types and some are disease specific. For instance, Cancer antigen 125, calretinin and vimentin are tumor markers that are expressed by tumor cells. Some of mesothelial cells markers are positive while some are negative. Markers like hyalurnon, phospholipids, protein phosphate inhibitors-1 are formed by mesothelial cells itself. Table below shows previously studied markers for mesothelial cells and their function.
TABLE:- Cell Markers for mesothelial cells
|
MARKER |
FUNCTION |
SOURCE |
REFERENCE |
|
AUA1 |
Immunocytochemical marker of epithelial cells |
Human |
[22] |
|
Calretinin |
Diagnostic tool for Hirschsprung Positive marker of mesothelioma |
Human |
[23][72] |
|
Cancer Antigen(CA) 125 |
Tumor marker Specific marker for mesothelial cell mass |
Human |
[24][73] |
|
CD44 |
Cancer stem cell marker |
Human |
[25] |
|
CD44H |
Receptor for hyaluronic acid Positive mesothelial marker |
Human |
[26][74] |
|
Cytokeratins |
Epithelial specific marker Expressed by mesothelial cells |
Human Rat |
[27][75] |
|
Cytokeratin 5/6 |
Mesothelioma marker |
Human |
[28] |
|
D2-40 |
Potential diagnostic marker of mesothelioma |
Human |
[29] |
|
Desmin |
Promising differentiating marker of reactive mesothelium from malignant epithelial |
Human |
[30] |
|
E-Cadherin |
Cell- cell adhesion marker Used for differentiating benign reactive mesothelial cells |
Human |
[31] |
|
HBME-1 |
Negative mesothelial marker in pleura Sensitive mesothelial marker in pleura |
Human Human |
[32][33] |
|
Hyalurnon |
Formed by Mesothelial cells Positive mesothelial peritoneal marker |
Human |
[34] |
|
Keratin |
Present in extreme amount in mesothelial cells Low amount or negative in mesothelioma |
Human Human |
[35] [36] |
|
Keratin 7 (K7) |
Highly expressed by mesothelial cells |
Human |
[37] |
|
Mcp130 |
Diagnostic tool for mesothelioma Also used for ovarian carcinomas |
Human Rat |
[38] |
|
ME1/ME2 |
Used for identifying both mesothelioma and normal mesothelial cells |
Human |
[22] |
|
Mesothelin |
Expressed by both reactive mesothelial cells and epithelioid mesothelioma |
Human |
[39] |
|
N-Cadherin |
Human mesenchymal stem cell marker Able to differentiate cardiomyocyte |
Human Human |
[30] [76] |
|
Phospholipids |
Formed by mesothelial cells Also known as positive marker for mesothelial cells in peritoneal fluid |
Human Human |
[34] [24] |
|
ProteinPhosphate Inhibitor-1 |
Consider as novel positive marker of mesothelium |
Mouse |
[40] |
|
Thrombomodulin |
Positive, specific and sensitive marker for mesothelial cells. |
Human |
[28] |
|
Vimentin |
Help in identifying mesothelial origin cells |
Human |
[41] |
|
Wilms Tumour 1 (Wt1) |
Marker used for differentiate between benign and epithelioid mesothelium Regulates transition of mesothelial into myofibroblasts |
Human Human |
[42] [77] |
2.0 Fibroblasts in peritoneum
To amend injuries in organ and tissues, cell response it in form of fibrosis. Resident fibroblast produces extracellular matrix and collagen. Myofibroblasts are the cells that are produced for carrying out the process of wound healing. In general, myofibroblast are gone through the process of apoptosis after completing healing process. It has been suggested that they become persistence after its mechanism fails and leading to more production and contraction of Extracellular matrix (ECM) and collagen causing fibrotic diseases.
2.1 Myofibroblasts in fibrosis
Myofibroblasts are defined as specialized subset of cells, which are considered as the key cells in tissue fibrosis. [9] Electron microscopy showed that myofibroblast have disorganized membrane (changes that occur in actin cytoskeleton), showed presence of stress fibers, microfilament bundles and also showed adhesions in the fibroblastic cells of granulation tissue. [10] Further electron microscopy showed the presence of gap junctions which connects myofibroblasts, thus strengthening the idea of resemblance between myofibroblasts and smooth muscle cell. Myofibroblast main functions are wound healing and connective tissue remodeling.
2.2 Fibroblast differentiation into Myofibroblast
To study and analyze functions of myofibroblast, a series of both in-vivo and in-vitro experiments were carried out by Hinz and Gabbiani. [15] They defined the origin of myofibroblasts as a result of 2 step differentiation of fibroblast process. [78] In initial step, fibroblasts differentiate into proto-myofibroblasts during mechanical stress and results in production of stress fibers which contains cytoplasmic actin. They also function to express and establish cellular fibronectin and are also enable to produce contractile force. Term “proto-myofibroblast” was given to these activated fibroblasts to differentiate them from inactive fibroblasts which are unable to produce contractile force. TGF-β1 along with ED-A fibronectin, under mechanical stress boosts further differentiation of proto-myofibroblast into myofibroblast. They are categorized by extensive α-smooth muscle actin level of expression and produces stronger contractile force than proto-myofibroblasts. This is also contributing into connective tissue remodeling. Gabbiani and Hinz also stated that unlike dermal wound healing, in pathological wound healing myofibroblasts do not vanish through apoptosis instead they lead to tissue deformation. In fact, contractures triggered by them are recognized characteristic of organ fibrosis. [79]

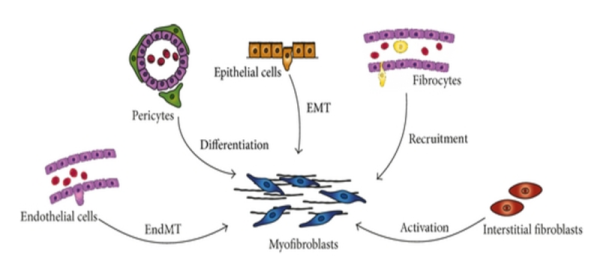
2.3 Hypothesis surrounding Myofibroblasts origin
The origin of Myofibroblasts in fibrotic tissue has been much debated, with disparate findings likely to stem from the range of different fibrotic diseases investigated and the array of technical approaches taken. [11] Most of the proposed theories have different cellular sources and mechanisms which lead to their origin (Figure 3).
Previous studies on dermal fibrosis proposed that mesenchymal-like cells known as pericytes are resident source of myofibroblasts. Immunohistochemistry and double immunofluorescence of patients’ biopsies showed phenotype link between pericytes and myofibroblasts. [80] It has been described that pericytes can differentiate into osteoblast, adipocytes, and smooth muscle cells. Research carried out by Lin et al., on Col1a1-GFP mice and Humphreys et al., using FoxD1-Cre knock-in mice, both transgenic mice with aim to identify pericytes in genetically labelled renal stroma to suggest that major source of myofibroblasts is pericytes, a contractile cell. [12, 13] A limitation of the aforementioned studies is the lack of lineage tracing strategies and the use of non-specific markers to identify pericytes. Markers such as CD73, PDGFR- β (Platelet-derived growth factor receptor- β) and neural/glial antigen 2 (NG2) used in these studies are also expressed by resident fibroblasts.
Endothelial to mesenchymal transition (EndoMT), similar process to EMT can be another potential source myofibroblasts. Study done by Zeisberg on three independent mouse models with chronic kidney diseases to study renal fibrosis found that EndoMT may contribute in fibrosis as about 30-50% myofibroblasts were able to express α-SMA and fibroblast-specific protein-1(FSP-1) and also endothelial cell markers CD31. [16] This indicates myofibroblasts may originate from endothelial cells. Myofibroblasts may also be derived from bone marrow. An elaborate series of studies carried out by Direkze et al., used molecular staining techniques for the detection of Y chromosome markers in irradiated female mice which were transplanted with male bone marrow. [18] They used skin wounding and paracetamol administration to create selection pressure to encourage engraftment of bone marrow derived myofibroblasts. They were able to identify myofibroblasts in the gut through their ability to express of alpha- smooth muscle actin and their morphologies. These findings are controversial however, as other groups have failed to identify BM derived myofibroblasts.
A recent study carried out by Yi Ting Chen et al., did genetic fate mapping of Wilms Tumor-1 positive cells on peritoneal fibrosis models. Fibrosis was induced either by dialysis solution, overexpression of TGF-β1, or sodium hypochlorite. [19] Results of this study postures as a challenge to the significance of mesothelial cells as probable source of myofibroblasts. It showed rather than mesothelial cells, underlying submesothelial resident fibroblast give rise to collagen producing myofibroblast. Results of genetic fate mapping of WT1+ mesothelial cells showed that after peritoneal injury, remaining mesothelial cells repair the disrupted peritoneal membrane by contributing to remesothelialization, but do not undergo MMT. While previous studies used Wt-1 as a biomarker and Cre recombinase to trace the fate of mesothelial cells. This makes it obvious that Chen finding still needs to be confirmed. This study also The last proposed theory for origin of myofibroblast is epithelial-mesenchymal transition.
Epithelial-Mesenchymal transition (EMT) is commonly defined as process during which transition to mesenchymal cells, epithelial cells lose their cell-cell adhesion and their polarity. Followed by obtaining mesenchymal markers like FSP 1, HSP47, α-SMA and vimentin but loses epithelial markers. [20] Myofibroblasts derived from epithelial cells through EMT proposed theory got strong piece of evidence from immunohistochemical study of peritoneal fibroblasts of continuous ambulatory peritoneal dialysis (CAPD) patients carried out by J-Heffernan et al. [14] Biopsies samples were taken from CAPD patients and divided into four groups: normal (I), non CAPD uremic (II), CAPD uremic (III) and non- renal patients with inguinal hernia (IV). Markers like desmin, E-cadherin, cytokeratins, α-SMA and also CD34 were analysed for expression by mesothelial cells and resident fibroblasts. Group I and II did expressed CD34 but no other markers. The reverse pattern was observed in other two groups. Degree of peritoneal fibrosis is connected with loss of CD34 expression. Myofibroblasts appear to originate from both by activations of resident fibroblasts and by mesothelial-to-mesenchymal transition of mesothelial cells.
3.0 Mesothelial-Mesenchymal Transition (MMT)
Under regular conditions, Mesothelial-to-mesenchymal transition of mesothelial cells is an autoregulated biological process which functions to repair tissues but under uncontrolled situations (Long term Peritoneal dialysis) they can cause Peritoneal fibrosis.
3.1 Stages of MMT
Mesothelial -Mesenchymal Transition (MMT) is similar process to Epithelial – Mesenchymal Transition (EMT). Defined as a step-wise and complex process where mesothelial cells undergo modifications in their architecture and molecular functions. [21] [48] Simple steps in which process of MMT occurs starts with the downregulation of intracellular adhesion molecules causing loss of microvilli, apical-basal polarity and disrupting intracellular tight junction. These transition undergoing cells adopt front to back polarity, have increased in their migration capacity, morphological changes and are able to express α-SMA. The main objective of this process is to repair wounded tissues and also play role in process like migration of cells, inducing neoangiogenesis and production of extracellular matrix.
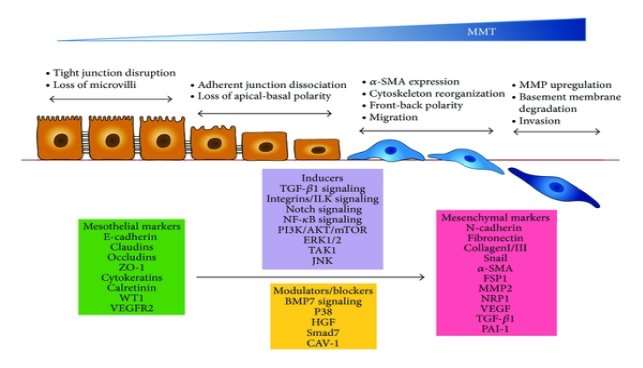

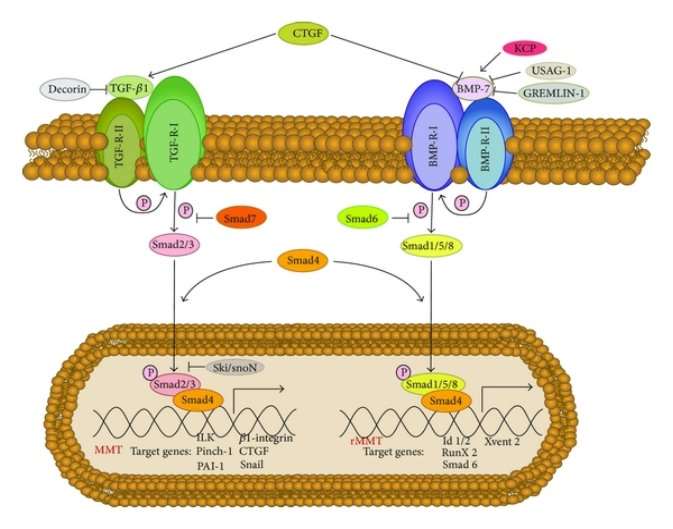
Molecules and signaling pathways that play role of either inducers or blocker during the process of MMT are TGF-β1 induced signaling pathways and BMP-7 signaling pathway. They are member of superfamily of growth factors, along with activins/inhibin and growth and differentiation factors (GDFs). Main attention is on growth factors because equilibrium between these factors maintains the epithelial like-phenotypic characteristics of Mesothelial cells and also in acquiring mesenchymal cells characteristics.
Studies not only showed that high levels of cyclooxygenase (COX)-2, CTGF and VEGF are expressed by mesothelial cells that undergo MMT. But also that there is connection between expression of these molecules and rate of transport of solute across peritoneal membrane. These molecules are also caught up into fibrotic, angiogenic and also in inflammatory responses. [49][50][51]
3.2 Cell Signaling Pathways
Activation of TGF- β, master molecule in peritoneal fibrosis during Peritoneal Dialysis, starts MMT signalling cascade with activating receptors that triggers smads- dependent (GSK-3, β-catenin, Lef-1/Tcf, integrin-linked kinase) and smads-independent signalling (H-Ras/Raf/ERK) pathway. [48][52][53][54]
Smad are intracellular proteins that functions to transfer signals from TGF-β to nucleus for gene transcription. Smad dependent signalling pathways activates when TGF-β and BMP-7 binds with their respective receptor type II (TGF R-II and BMP R-II respectively). The binding leads to activation of receptors type I of TGF and BMP-7. These receptors type I functions to carry out phosphorylation process. TGF R-I phosphorylates Smad2 and Smad3 in TGF-β pathway and Smad1, Smad5 and Smad8 by BMP R-I. These activated Smads forms heterodimers with Smad4. Followed by translocation of these complexes into the nucleus where they function to activate genes involved in either process of MMT and rMMT (Fig. 5) Presence of few other BMP-7 modulators may affect the signalling of BMP-7. Smad dependent signalling pathway of TGF-β and BMP-7 are controlled by Smad7 and Smad6 respectively.

In case of Smad independent pathway of TGF-β and BMP-7 involves a protein kinase TAK1, which functions to regulate processes like transcription and apoptosis. In TGF-β signalling pathway, TRAF6 initiates activation of TAK1 which further leads to activation of p38 and MAPK-JNK signalling pathway. Receptor type I of TGF-β phosphorylates Shc, which helps in activating MAP kinase ERK1/2 signalling pathway. p38 pathway act as blocker of mesothelial cells conversion to mesenchymal. Maybe it’s due to the fact that XIAB, a protein belonging to the family of inhibitor of apoptosis protein helps BMP-7 to activate p38 signalling pathway. TGF-β also encourages PI3-K/Akt pathway by inactivating GSK-3β, to stabilize β-catenin and snail and to activate mTOR. This results in localisation of β-catenin into the nucleus, where they interact with LEF1/TCF, in order to feed into Wnt signalling pathway. They also contribute in the transcription of mesenchymal-related genes. Furthermore, localisation of snail into the nuclear encourages transcriptional repression of E-cadherin and several other adhesion molecules.
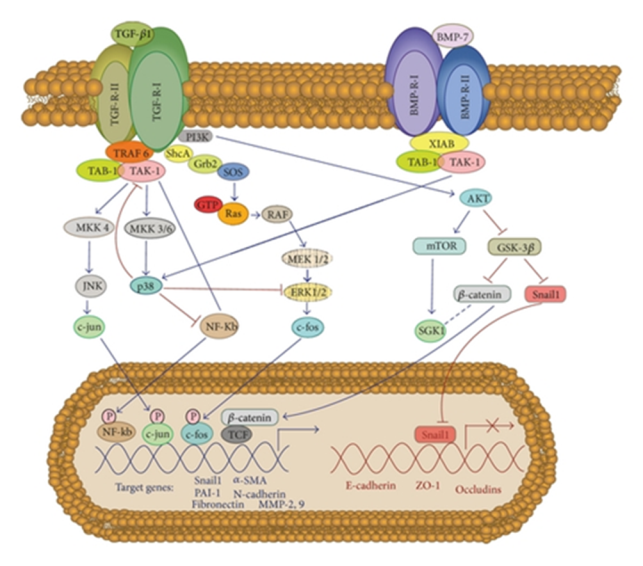

Smad dependent or independent pathway, both depicts the fact that TGF-β plays a key role in developing fibrosis by promoting mesothelial to mesenchymal transition. Another fact is BMP-7 plays role of antagonist to TGF-β. Utmost studies has used TGF-β1 as their target goal to design therapeutic treatment for peritoneal fibrosis. Most of them intend to control TGF-β1 activity either via drug or synthetic proteins design specifically for this purpose.
3.3 Evidences of Mesothelial-mesenchymal transition (MMT)
There are more than enough studies results which clearly supports the process of MMT occurring fibrosis but in 2003, Yanez-Mo et al., published breakthrough in-vitro and in-vivo study where they were able to demonstrated MCs undergoes mesenchymal conversion. [46]. Primarily in vitro studies were carried on by isolating mesothelial cells from patients undergoing peritoneal dialysis. These isolated cells were characterized using immunostaining, western blotting, flow cytometry and reverse-transcriptase polymerase chain reaction. Mesothelial cells from omentum from healthy persons were used as control and all results were compared. Morphological analysis sample showed varies morphologies ranging from cobblestone like appearing cells to fibroblast like cells. Mixed morphology and non-epithelioid were also observed in few samples. Level of E-cadherin, cytokeratin and vimentin were analyzed for the mesothelial-to-mesenchymal transition in mesothelial cell cultures. Results demonstrated noticeably low level of E-cadherin and cytokeratin expression in both cobblestones like cells and non-epithelioid cell as compared to omentum cell cultures. In non-epithelioid cells, vimentin has high expression level. Expression of snail and E-cadherin in mesothelial cells isolated from effluent and omentum were analyzed by the process of RT-PCR. While in omentum mesothelial cells, showed no presence of snail mRNA but increase in expression of these messenger RNA in effluent derived mesothelial cells. These results were established after carrying out in vivo studies of peritoneal biopsy samples.
With dual aim of studying anatomical changes in peritoneal membrane and correlating them with functional parameter, Del Peso and team carried out a clinical study. [47] They used peritoneum biopsies taken from thirty-five steady patients which were still taking peritoneum dialysis treatment form last 2 years. They observe loss of mesothelial layer in about 74% patients, fibrosis in 46%, 17% showed presence vasculopathy. With submesothelial cytokeratin staining, 17% showed evidence of EMT. Interestingly, they found that myofibroblasts were present in all patients with EMT but only in 36% without EMT.
3.4 Potential therapeutic approaches to reverse MMT (rMMT)
Several studies describe therapeutic strategies which are targeting anti-EMT of Mesothelial cells are emerging as promising treatment against fibrosis development. Tamoxifen is a drug used for treating breast cancer. The tamoxifen is oestrogen receptor and their activity affects action of TGF- β1 has been reported. In year 1991, Clark and team showed noticeable decrease in level of peritoneal fibrosis, when given orally to peritoneal fibrosis patients.
Loureiro and team also used this drug to study its effect in early stage of PD induced peritoneal fibrosis. [61] Theyused PD model of mice for in-vivo study and along with that they also did in-vitro study. In vitro studies were carried out with mesothelial cells (MCs) derived from omentum and effluent. Mesothelial cell sample cultures were checked for any change in morphology and also level of E-cadherin expression after treating them with Tamoxifen. For effluent derived MCs, MMT was not reversed by Tamoxifen but some mesenchymal molecules expression was reduced. In mice PD model, they confirmed that MMT and thickness of peritoneal membrane increases parallel. They also observed that Tamoxifen does reduce thickness of peritoneal membrane and improves its function.
They also administrated P17 and P144, synthetic peptides that were designed to bind with TGF-β1 directly. [62] It has been reported that in organs like liver, heart, skin these peptides have shown strong antifibrotic effect by blocking biological function of TGF-β1.
Bone morphogenic protein-7 (BMP-7) and Hepatocyte growth factor (HGF), are two molecules that were successfully demonstrated their ability to inhibit and reverse process of Mesothelial to mesenchymal transition and renal fibrosis in animal models by X Y D et al., and Yang et al. [64] [65]
In vitro study carried by Yang and team, where they used histone acetyltransferase inhibitor (HAT) C646 in EMT model of HPMCs. [66] EMT was induced by treating with high glucose. C646 treatment of EMT model, found TGF-β1/Smad3 signalling was blocked. This might provide with a novel therapeutic treatment for peritoneal fibrosis.
Even though most of the therapeutic studies focused on mesothelial mesenchymal transition process and TGF-β1, there are other factors that contributes to loss of functional properties of peritoneum. Thus promotin
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


DMCA / Removal Request
If you are the original writer of this essay and no longer wish to have your work published on UKEssays.com then please click the following link to email our support team:
Request essay removal