Theories of Colour Vision
| ✅ Paper Type: Free Essay | ✅ Subject: Sciences |
| ✅ Wordcount: 2454 words | ✅ Published: 08 Aug 2018 |
- Kishan Lakhani
How does colour vision work?
It is difficult to imagine a world without colour perception as it is constantly in action by a whole spectrum of living organisms and for a range of purposes, it “not only allows us to detect objects that might otherwise be obscured by their surroundings; it also helps us to recognize and identify things we can see easily” (Goldstein, 2000, p.203), thus making it an essential component of vision. Colour dictates survival in many environments; the artic fox boasts a white pelt allowing effective camouflage over its prey and a significant factor in its ability to hunt (Sekuler & Blake, 2006). I will explore the mechanisms that are said to explain colour vision at the photoreceptor level and beyond, but first of all we must understand what colour actually is.
The electromagnetic spectrum ranges from cosmic rays to radio waves, as wavelength increases. Between UV and Infrared lies a strip, visible light, this is what we are interested in with regard to colour (Snowden et al., 2006). Colour arises when light rays from this visible spectrum (390nm-750nm) are reflected off objects and into our eyes. Differing wavelengths of light from this spectrum subsequently determine the colour perceived, as short wavelengths produce the colour violet (350nm), medium and long wavelengths produce green and red respectively. Sir Isaac Newton famously wrote “The rays to speak properly are not coloured. In them there is nothing else than a certain Power and Disposition to stir up a Sensation of this or that colour” (Sekuler & Blake, 2006, p.236) So colour doesn’t really exist in the physical world at all, it is our own psychology that creates the concept of colour. Semi Zeki (1983) refined Newton’s words further saying that colour vision is a “property of the brain, not the world outside.”( Sekuler & Blake, 2006, p.236) Hence we can sum up that colour vision lies in the eyes and brain – and not in the physics of light itself (Anderson 2012). So to answer the question ‘How does colour vision work’ we must explore the visual systems of the organisms further.
Figure 1 The Electromagnetic spectrum (2012), Diagram showing the visible spectrum within the electromagnetic spectrum

Photoreceptors are light sensitive cells found at the back of the eye in the retina. They contain visual pigments that absorb photons of light and convert this light energy into chemical energy, this process is called phototransduction. Within the pigment is a protein which determines the wavelength of light absorbed by the pigment and also a chromophore which is responsible for absorbing photons of light. (Wolfe et al., 2009) There are 2 types of photoreceptors, rods and cones. Cones are mainly concentrated in the fovea in the centre of the retina whereas Rods are completely absent in the fovea and more prevalent in the periphery of the retina (Snowdon et al,. 2006). Rods are adapted so they can operate in low light levels due to spatial pooling allowing scoptic vision, whilst sacrificing visual acuity. They contain just one pigment rhodopsin – denying colour vision due to the univariance principle. Cones however contain 3 photo pigments, which are sensitive to various wavelengths of light and can therefore let us see colour.
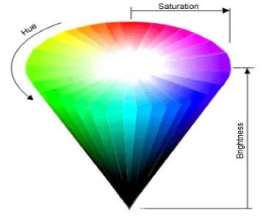

In comparison to the physical description of light, colour is much easier to describe as it is experienced psychologically not physically – such is the nature of light. It can be specified by just three values; hue, saturation and lightness.(Palmer, 1999) This is very important with reference to colour vision as it implies that many different lights will produce the same colour experience (Palmer, 1999). The three perceptual dimensions of colour can be summarised in what is known as the colour spindle. Hue is referred to as “the chromatic aspect of light” (Wolfe et al., 2009) and is dictated by its wavelength. Saturation corresponds purity and how vivid the colour is (Palmer 1999), and brightness involves the intensity of the colour (physically).
The Young/Helmhotz trichromatic theory of colour vision works on the photoreceptor level, and ultimately was sprouted through the results of Helmhotz’s colour matching experiment. Observers were allowed to vary the intensities of 3 primary lights and mix them (in a comparison field) to match the colour of a single wavelength in a test field. They had to find a psychological match between the mixture of primary lights and the test light, simply by varying the intensities of the primary lights. This match is known as a metameric one, as the light in the comparison field is physically different yet psychologically identical to that in the test field. Results showed that by varying intensities in the comparison field, the observer could find a metameric match using just three primary lights. Dichromatic observers were unable to find matches for every colour in the test field. In summary, “with 3 primaries, you can get any combination of responses in the 3 cone types, so you can match the appearance of any test light.” (Anderson 2012) It is therefore clear to us that colour vision heavily relies on three different receptor mechanisms or cone photopigments, each with different spectral sensitivities (Goldstein, 2010). This is the basis of the trichromatic theory I will now elaborate on.
Monochromats possess only one type of pigment in their cones. In this case, the ability to see colour is not possible. The same chain of events is initiated in the visual receptor despite there being a variety of wavelengths in the light absorbed by the pigment. The receptors response conveys information about how much light has been absorbed, but this response provides no information about the wavelength of this absorbed light. (Sekuler & Blake, 2006) It is therefore impossible to discriminate wavelength when there is only one photopignment, which has uniform spectral sensitivity. The response could have altered due to a change in wavelength or light intensity, and monochromats are none the wiser – this is known as the principle of univariance. This situation is not unique to monochromats, as in low light levels rods are the only photoreceptors in action. They have one photopigment, rhodopsin resulting in the same dilemma. Consequently we “disregard the wavelength information and see an image that appears in shades of grey” (Snowden et al., 2006) explaining why we can’t monochromats can’t see colour and why none of us have colour vision in low light levels.
Dichromats possess two photopigments, which is very useful in terms of colour vision. The two pigment types have different absorption spectra, extracting some usable wavelength information about light (Sekuler & Blake, 2006) It is now possible to separate and disentangle wavelength and intensity, allowing colour to be visible to a certain extent. Certain wavelengths are confused and constitute failures of discrimination (Sekuler & Blake, 2006). A key reason that leads to the idea humans aren’t dichromats revolves around what is known as the neutral point. All dichromats possess this neutral point in which a single wavelength is always confused, and “the existence of a single neutral point is the hallmark of a two-pigment eye.” (Sekuler & Blake, 2006, p.249) As humans do not show any traits of having the neutral point, there is a strong belief that humans have more than two cone photopigments.
Trichromats hold three cone photopigments, enabling total discrimination of wavelengths throughout the visible spectrum. Trichromacy also ties in with Helmhotz’s colour matching experiment, indicating three not two photoreceptors are required for complete colour vision across the entire visible spectrum. The three pigments are most sensitive to light of a particular wavelength – approximately 430, 530 and 560 nanometres respectively. (Sekuler & Blake, 2006) Figure 4 shows the each cone pigment absorbs a very wide range of wavelengths. So between 400nm and 650nm there are at least two types of cone photopigments absorbing light. In the region of 475nm, all three types of pigments are affected and stimulated. (Sekuler & Blake, 2006) Hence we can conclude that the absorbance range is tremendously increased with three photopigments, and as light is reflected onto the retina every wavelength of light in the visible spectrum can be perceived in the form of colours by our brain.


Trichromatic theory may explain how the existence of just three cone photopigments allows colour matching for any wavelength of light in the visible spectrum, using just three primary lights. However, it still leaves many unanswered questions when it comes to having a full understanding of how colour vision works – as Hering highlighted. Negative afterimages, the visibility of four psychologically pure hues (blue, red, green and yellow) and the absence of complementary hues such as blueish – yellow all indicate that trichromatic theory alone is inadequate in explaining how colour vision works.


Hering made an important discovery with regard to afterimages. If we stare at the black dot in Figure 5 for around 30 seconds, and then look at a blank piece of paper we notice the colour of each square changes. The green changes to red and red to green, whilst the blue changes to yellow and the yellow to blue. Based on results like these, Hering proposed the concept that red and green are paired and blue and yellow are likewise paired. (Goldstein, 2010) An experiment where observers were shown patches of colour, and then asked to estimate the ratios of blue, green, red and yellow from each patch they received. Results showed that observers very rarely saw blue and yellow, or red and green together. (Abraham & Gordon, 1994 cited in Goldstein, 2010) Sekuler and Blake (2006, p.255) also support this view that these complementary hues do not coexist, as “an object never appears both blue and yellow at the same time.” Hurvich and Jameson’s hue cancellation experiment strengthens the case further, as any redness was eliminated when a green light was added to the red light. Hering also observed that those who are colour blind to red, are also colour blind to green; which ultimately led him to declare the opponent-process theory of colour vision. (Goldstein, 2010) So we can deduce from this evidence that blue is paired with yellow and red with green; the basis of the opponent-process theory.
The opponent-process theory of colour vision follows the trichromatic theory, rather than replacing it, the two work hand in hand to explain how colour vision works. In fact it is the responses from the cones of the retina that form the basis of the opponent channels. (Anderson 2012) There are three opponent channels, two chromatic and one achromatic, and are formed by combining the responses from the three cone types. (Sekuler & Blake, 2006) Figure 6 displays the red-green chromatic channel is comprised from the outputs of the M and L cones. It is also known as the M – L channel, as it signals the difference between the outputs of the M cones and of the L cones. (Sekuler & Blake, 2006) The second chromatic channel is the blue-yellow channel, and it represents the difference between the S cone outputs and the sum of the M and L cone outputs. (Sekuler & Blake, 2006) It is therefore also referred to as the S – (M + L) channel. The achromatic channel is known as the luminance channel, and combines the output of the M and L cones so we can also label it the M + L channel. The activity in this luminance channel hinges on the sum of excitation of both M and L cones (Sekuler & Blake, 2006). This addition can lead determine an object’s visibility, “The shape of the photopic sensitivity curve (closely related to visibility) can be predicted by taking a sum of M and L cone responses.” (Smith and Porkorny, 1975 cited in Werner et al., 1984).


Russel DeValois was responsible for the finding of opponent neurons in the retina and lateral geniculate nucleus (LGN), which could provide physiological evidence to back up Hering’s propositions. (Goldstein, 2010) The LGN is the station responsible for receiving input from the retina and transmitting it to the visual cortex. Devalois conducted experiments on LGN cells of monkeys (who have the same trichromatic vision as ours), and discovered opponent cells which behaved as if subtracting outputs from different cones and also nonopponent cells which behaved as if adding outputs from different cones. Devalois discovered opponent cells reproduced an ON or OFF response determined by the wavelength of light. (Sekuler & Blake, 2006) This can explain the first chromatic channel Hering proposed (M – L) channel. Long wavelength cone excitation results in a positive or ON response, whilst medium wavelength cone excitation results in a negative or OFF response. Hence if the net response is positive then a red colour is visualised (long wavelength of light), and similarly a blue colour is perceived if the net response is negative. This supports Hering’s initial observation that the hues red and green cannot coexist. Opponent cells were also responsible in explaining the S – (M + L) channel. Short wavelength cone excitation results in a positive or ON response, whilst wavelengths around 580nm (M+L) cone excitation results in a negative or OFF response. Further findings included the fact that nonopponent ON cells produced ON responses for every wavelength, although some wavelengths produced stronger responses than others and OFF cells produced OFF responses for every wavelength again with varying strengths. It is these nonopponent cells which form the achromatic channel outlined by Hering. (Sekuler & Blake, 2006)
All in all, colour vision begins at the photoreceptor level as explained by trichromatic theory. The outputs of the three cone photopigments have been redistributed into the achromatic and chromatic channels at the LGN, as trichromacy progresses to opponent-process theory. Palmer (1999) concludes by describing the dual process theory; in which the products from the trichromatic stage are used as the inputs for the secondary opponent-process stage. As we venture from the LGN, further into the visual system, the information is perceived by the visual cortex of the brain facilitating us with colour vision.
Bibliography
Anderson, S (2012). Colour vision, Vision and visual perception, Optometry. Aston University
Dimitri Poumidis, (2008), Spectral Sensetivities [ONLINE]. Available at: http://www.gravurexchange.com/gravurezine/0805-ezine/ploumidis.htm [Accessed 25 January 13].
Goldstein, E. B. (2010). Sensation and perception (8th ed.) Chapter 9. Wadsworth Cengage Learning
Joshua Stevens, Jennifer M. Smith, and Raechel A. Bianchetti , (2012), The Electromagnetic Spectrum [ONLINE]. Available at: https://www.e-education.psu.edu/geog160/node/1958 [Accessed 03 January 13].
Marc green, (2004), Opponent process theory [ONLINE]. Available at: http://www.visualexpert.com/FAQ/Part1/cfaqPart1.html [Accessed 09 February 13].Paul Schils , (2012), Chromatic adaptation [ONLINE]. Available at: http://www.color-theory-phenomena.nl/12.00.htm [Accessed 08 February 13].
Palmer, S. E. (1999). Vision science: photons to phenomenology, Chapter3. Massachusetts Institute of Technology
Sekuler R. & Blake R. (2005). Perception (5th ed.) Chapter 2. McGraw-Hill
Sekuler R. & Blake R. (2005). Perception (5th ed.) Chapter 7. McGraw-Hill
Snowden R., Thompson P. & Troscianko T. (2006). Basic Vision, Chapter 1. Oxford University Press
Snowden R., Thompson P. & Troscianko T. (2006). Basic Vision, Chapter 5. Oxford University Press
Tom Jewett, (2009), Hue, Saturation, Brightness [ONLINE]. Available at: http://www.tomjewett.com/colors/hsb.html [Accessed 10 January 13].
Wolfe, J.M., Kleunder, K.R., Levi D.M., et al (2009). Sensation and perception (2nd ed.), Chapter 5. Sinauer Associates Inc
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


DMCA / Removal Request
If you are the original writer of this essay and no longer wish to have your work published on UKEssays.com then please click the following link to email our support team:
Request essay removal