Quantification of Biofilms
| ✓ Paper Type: Free Assignment | ✓ Study Level: University / Undergraduate |
| ✓ Wordcount: 5844 words | ✓ Published: 06 Jun 2019 |
The ability of bacteria to easily History and Definition of biofilms
Although the free-floating, planktonic form of most bacterial species is the first known and widely studied in the laboratories, later studies have shown that microorganisms predominate naturally as surface attached matrix – enclosed populations (Costerton et al. 1995; Abelson and McLaughlin 2012). In their native habitat, microbes most often live in hydrated ultra-thin layers coating on surfaces. As they establish new habitats, their secretions construct an elaborate film-like meshwork that provides additional surface area for bacterial expansion. These coatings are called biofilms. Although the word biofilm was used many decades after the discovery of microbiology, several references (Heukelekian and Heller 1940; Henrici 1933; Zobell 1943) have described the aggregative life form of microorganisms surrounded by self-produced matrix. Anthony van Leeuwenhoek, the father of microbiology in his letters published from Sept 1683 to June 1708 describe the dental plaque and the mucous membrane of the tongue from his mouth as follows, ‘I saw an inconceivable number of exceeding small animalcules and these of divers sorts. But far the greatest number was of one and the same bigness, yet so little that they could not be discerned but by great attention, through a very good magnifying glass; and most of these animalcules were abiding where the said matter from the tongue lay, and I took into consideration whether the said creatures might not indeed be getting their food from the particles of the tongue.’ Louis Pasteur in the study of vinegar production by acetic acid fermentation used bacteria growing on the surface of chips of beech. Later references to this sessile life form of microorganisms were made in marine microbiology by Angst, 1922; Henrici, 1933; ZoBell and Allen, 1935. They used the terms slime and films to describe bacterial adhesion, aggregation and multiplication on surfaces (Høiby 2014).
Biofilms are archaic and an advantageous model of microbial growth. Although some authors claim that the term ‘biofilm’ was coined by Costerton in 1978 (Chandki, Banthia, and Banthia 2011) , it was first used in a publication in 1975 by Mack and Ackersonin an article titled ‘Microbial film development in a trickling filter (Mack, Mack, and Ackerson 1975). Biofilm was used to describe microbial growth and development. It was next employed by the Canadian microbiologist J.W. (Bill) Costerton in the early 1980s (Abelson and McLaughlin 2012; Høiby 2014). A biofilm is a complex of microorganisms associated with a biotic or abiotic surface and embedded into an extracellular matrix (ECM) of macromolecules with changes in their cellular physiology, representing a differential expression of genes (Fanning and Mitchell 2012). It can also be put as an assemblage of surface-associated microbial cells that is enclosed in an extracellular polymeric substance matrix separated by a network of open water channels; a well organised cooperating community of microorganisms(Kokare et al. 2009; Donlan 2002). According to (Donlan 2002) it is an assemblage of microbial cells that is irreversibly associated (not removed by gentle rinsing) in a complex 3-D gelatinous matrix of extracellular polymeric substances generated by the microbial cell (Kokare et al. 2009). It is a city of microbes (Watnick and Kolter 2000) associated with extra polymeric substances which are known to confer specialized functions and behaviour often not found in the non-biofilm forming organisms.
Biofilms are not only found on abiotic surfaces but in living tissues such as inner ear, heart valve, lungs, etc. or secretions and components of its environment may be found in it. Biofilms are the principal form of microbial growth and are critical to the development of clinical infection and fermentation. In nature, biofilms have high levels of organisation and may exist as single or multiple species communities in a single layer. The structure of biofilms is an optimal environment for cell-cell interactions, including the intercellular exchange of genetic material, communication signals, and metabolites, which enables diffusion of necessary nutrients to the biofilm community and removal of waste products (http://www.sysbio.org/sysbio/biofilms.stm, (Nadell et al. 2015). The matrix in which microbes in a biofilm are embedded protects them from UV exposure, metal toxicity, acid exposure, dehydration and salinity, phagocytosis, antibiotics and antimicrobials. Microbial cells within biofilms are in orders of magnitudes more resistant to stress than free floating cells.
The structure of biofilms
Biofilms are highly organised community of microorganism. Bacterial biofilms are composed mainly of microbial cells, extracellular polysaccharides, DNA, water, lipids and proteins (Rabin et al. 2015; Vasudevan 2014) as shown in Table 1. This assemblage is defined in structure and composition. The exopolysaccharides form the backbone to which the other components bind to and it accounts for 50-90% of their total organic carbon (Vasudevan 2014). Within the biofilms are microcolonies which are separated by water channels to allow the flow of nutrients, oxygen, water and microorganisms from one site to another.
Table 1. Composition of biofilms
| S/N | Component | Percentage of matrix |
| 1 | Water | Up to 97% |
| 2 | Microbial cells | 2-5% |
| 3 | Polysaccharides | 1-2% |
| 4 | Proteins | <1-2% (includes enzymes) |
| 5 | DNA and RNA | <1-2% |
| 6 | Ions | Bound and free |
(Vasudevan 2014)
The exopolysaccharides secreted are synthesized extracellularly or intracellularly and secreted by cell-wall anchored enzymes into the outside environment where they serve as anchors for other carbohydrates, proteins, nucleic acids and lipids to adhere. Due to the high moisture content of the EPS, hydrogen bonding between the microbial cells is favoured. The bacterial polysaccharide in Gram negative and Gram positive bacteria are polyanionic or neutral and cationic respectively (Vasudevan 2014). The extra polymeric substances of most bacteria are comprised of hexoses. An analysis of exopolysaccharide composition and linkages of a group of organisms (Enterococcus faecalis, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp) termed ‘‘ESKAPE’’ due to their ability to escape treatment with commonly used antimicrobials. (Rice 2008) showed that although these carbohydrates are not unique to biofilms, they differ from one organism to another. Exopolysaccharides of all the strains have high total contents of mannose followed by galactose and glucose as the second and the third most abundant carbohydrate in all the strains. N-acetyl-glucosamine , galacturonic acid, arabinose, fucose, rhamnose and xylose are less common with-acetyl-glucosamine , galacturonic acid occurring in 6 out of the eight strains and arabinose, fucose, rhamnose and xylose in 5 strains (Rabin et al. 2015; Bales et al. 2013). For the linkages, in agreement with the glycosyl arrangement, a significant degree of the aggregate linkages included mannose.
The proteins in the extracellular polymeric matrix which in some cases as in activated sludge flakes exceed carbohydrates have not been studied as extensively as the exopolysaccharides. There are many proteins associated with biofilms. They can be broadly grouped as extracellular (e.g. aminopeptidase),cytoplasmic (e.g. ornithine-carbamoyl transferase), periplasmic (e.g.TolB), membrane (e.g.porin proteins) and outer membrane associated proteins (Zhurina et al. 2014).
It is important to note that the makeup and organisation of EPS among various microorganisms is heterogeneous and this variety may be attributed to many elements such as culture, development stage, process parameter, bioreactor sort, extraction strategy, and scientific tools used (Sheng, Yu, and Li 2010).
Biofilm formation
Biofilm formation is a complicated process that involves a sequence of steps such as the formation of conditioning layer, bacterial adhesion, bacterial growth, biofilm maturation and dispersal as shown in Figure 1.
Conditioning and adhesion
The substratum and the microorganisms are essential for biofilm formation and the absence of one inevitably means no biofilm formation (Garrett, Bhakoo, and Zhang 2008). Without a surface, there are a very few bacteria and biofilms are highly favoured in solid -liquid interface. The nature of the surface otherwise known as the conditioning of the surface has a lot of influence on the attachment of microorganism and it is determined by the inorganic and organic materials in the medium which settle on the available surface by gravitational force (Garrett, Bhakoo, and Zhang 2008). This alters the original properties such as hydrophobicity, electrostatic forces, etc of the surface thus providing platform and nutrients to favour bacterial growth. Most environmental surfaces possess negative charges which repel the negative charges on the bacterial surface. However, as the layering of the surface builds up, it conditions the surface to acquire a charge and it becomes increasingly attractive to bacteria carrying an opposite charge.
The characteristics of the surface of substratum and that of the microorganism determine biofilm formation. Microbial colonization increases with rough surfaces as shear forces are reduced and more surface area are available to bacterial attachment (Donlan 2002). The physiochemical properties of a surface also has a large influence on the rate and extent of bacterial attachment. Microorganism appear to attach faster to hydrophobic nonpolar surfaces like Teflon and plastics than to hydrophilic surfaces like glass and metals (Donlan 2002).
The trigger for planktonic cells to become sessile is often determined by the wettability and the free charges on the surfaces of the bacteria and the substratum. The rate of attachment is influenced by several factors such as the force of attraction between the bacterial cell and the surface and the nature of the surface. The proportion of the surface affects the rate of growth of the bacteria and this was demonstrated in early studies on bacteria growth by ZoBell and Anderson in 1936 and Lloyd in 1937 (Zobell 1943). They showed that in dilute solutions, smaller vessels presented more solid surface per unit of water which encourages bacterial growth. The characteristics of the substratum affect biofilm formation and it is generally accepted that the rougher and more hydrophobic a substrate is, the quicker a biofilm to develop on it (Donlan 2001).
The first stage of biofilm formation which is adhesion is usually in two stages: reversible and non-reversible stages. Reversible adhesion occurs within a few second of bacteria making contact with the substratum and According to Derjaguin–Landau–Verwey–Overbeek (DLVO) theory [34], there are forces which come into play for initial adhesion and they are namely, van der Waals attractive forces (vdW), steric interactions and electrostatic repulsive. DVLO theory summarises the interactive forces between colloids and flat surfaces using the balance between two opposing forces — Electrostatic repulsion and Van-der Waals attraction.
The electrostatic forces between the bacterial surface and the surface together with the use of flagella and pili (type IV) provide mechanical attachment to the surface. Flagella and fimbriae are essential for the initial interactions between the bacteria and surface and cell aggregation into micro colonies respectively (Rabin et al. 2015). The electrostatic forces called van der Waals forces are weak and reversible at this stage and the microorganisms are easily removed and killed by mild antimicrobials and/or hydrodynamic forces.
Irreversible attachment occurs when an organism has attached firmly and steadfastly on a surface and can withstand shear stress. The set of genes transcribed in biofilms are different from those copied in planktonic life forms and genetic studies show that surface contact simulates different genetic expression of factors that favour sessility such as those involved in biofilm formation in response (Kostakioti, Hadjifrangiskou, and Hultgren 2013). The variation in genetic expression of the bacteria at this stage is attributed to quorum sensing. Quorum sensing is a cell density dependent inter cell communication medium via inducers to induce or repress the expression of genes involved in biofilm maturation and detachment. The attached cells begin to express surface appendages such ‘as pilli, flagellum and extracellular polymeric substances. Firm irreversible attachment is attained with the aid of adhesive appendages such as type 1 pili for uropathogenic Escherichia coli (UPEC)and other E. coli pathotypes(Martinez et al. 2000); fimbriae in P.aeruginosa (O’toole and Kolter 1998)and by mean of adhesins and pili for Enterococci (Kafil and Mobarez 2015). EPS is a key player in the transition from reversible to a non-reversible attachment and strong adhesion of cells (Janissen et al. 2015).
Non-reversible adhesion occurs within minutes of attachment and here a different set of forces come into play. These forces include dipole-dipole interaction, hydrogen bonds, hydrophobic, and ionic covalent bonding
BACTERIA IN HARSH ENVIRONMENTS
Microorganisms can be found almost in all environments even the most adverse of them like deserts, hot springs, salty lakes. The question that comes to mind is how they survive in these environments. There are many studied mechanisms by which bacteria survive and sometimes even thrive in these conditions. Some of these conditions are discussed below.
Bacteria are known to secrete cold and heat shock proteins which help them at low and high temperatures.
Bacteria also modify their cell walls
They can also DNA repair mechanism. The ability of bacteria to easily repair their DNA help them to quicky recover fron damaged to their cell components and this may the main cause of antimicrobial resistance.
Formation of biofilms
On dry surfaces microorganisms become attached to the surface and the physiology of attached cells is different from planktonic cells. They are often less susceptible to antimicrobials than planktonic cells but more susceptible than biofilms. Vickery et al alerted the world with findings that surfaces in the hospital have loads of microorganism and biofilms even with the routine cleaning and long period of time. By real time PCR, aerobic culture and SEM the team demonstrate the microorganism survive as biofilms over long period of time. From 44 sample collected from intensive care beddings, environments and hard surfaces, 41 of them showed biofilms by SEM. These biofilms were mostly polycolonials and consisted mainly of S. aureus (>50%) by PCR and by aerobic cuture methicillin-resistant S aureus (18%), vancomycin-resistant Enterococcus (8%) and extended-spectrum beta-lactamase Sphingomonas paucimobilis(11%). Live/dead stain showed unusual result of live cells on the sampled surfaces 12 months after samples were collected and stored in sterile cool environments. The ability for these e microorganism to survive for this length of time is quite unexpected and may answer to why there are many cases of nosocomial infections (65%) despite routine cleaning of hospital surfaces. It is noteworthy to say that sampled surfaces have been cleaned or treated with using detergent 500 ppm chlorine before they were sampled. Vickery suggested that there is a possibility the content if the antimicrobials may serve as a source of organic nutrients for these microorganisms in these biofilms. This changes a lot about popular ideas of routine cleaning in the health care setting and shows there is still a huge gap between curbing healthcare associated infections and the current regime of hospital cleaning. There are still a lot questions to be answered as to why microorganisms survive for long on dry surfaces.
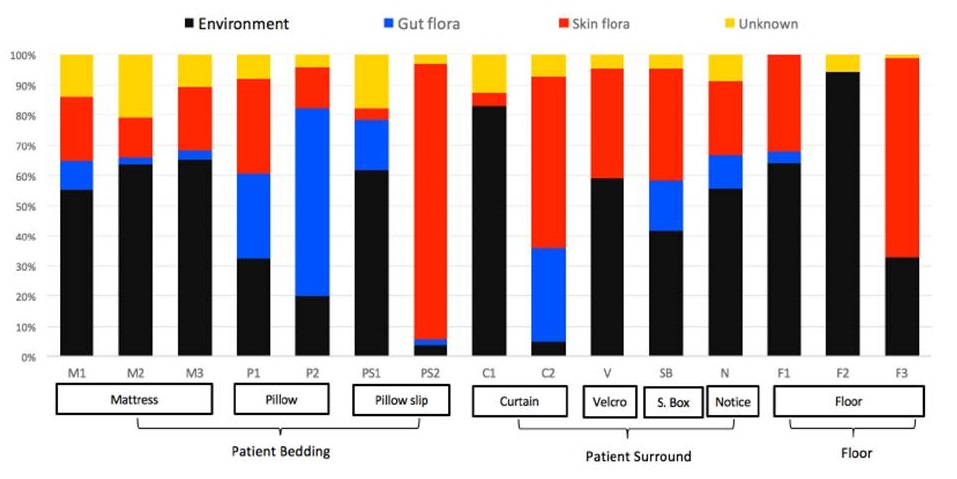
Figure 3: Composition of biofilms by bacterial source
Otter (2015https://reflectionsipc.com/2015/06/30/biofilms-make-the-hospital-environment-far-from-inanimate/
Rayar (2017) compiled the results generated from four countries on the isolation of multidrug resistant microorganisms (MDRO) biofilms on hospital surfaces. The results taken from the countries understudied which are United Kingdom, Saudi Arabia, Brazil and Australia showed that dry biofilms on dry surfaces is a worldwide problem. Samples were taken from intensive care units after recommended routine cleaning had taken place and analysed used sonication in tryptic soy broth for total viable counts of MDRO and assayed for biofilms by SEM and Live/ dead stain in confocal scanning electron microscopy (CFSM).
Very high percentage recoveries were made from the samples taken. From 40 samples taken from Brazil over 20 recorded MDRO and 100% of them were biofilm positive , in Scotland 4 out of the 8 sample had MDRO AND 68% biofilm positive , from the 28 samples taken from intensive care in Australia 20 were MRDO positive with 78% biofilm positive. Sample from Saudi Arabia were however not biofilm positive even though 13 out of the 20 sample taken were MDRO positive.
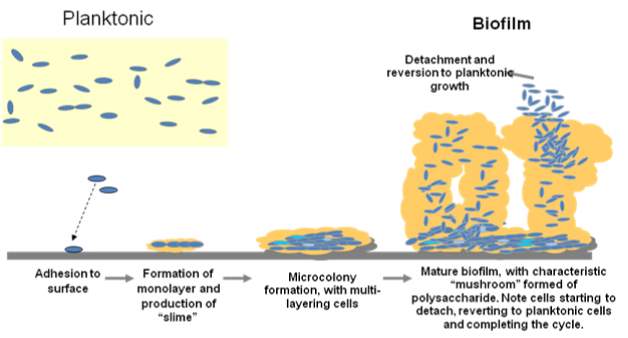
Fig 1: Various stages of biofilm formation and development
(Vasudevan 2014)
As the attached cells divide an initial layer of micro colonies is built. The cells continue to multiply upwards and outwards forming multilayers of colonies embedded in extracellular polymers. Matured biofilms are shaped like mushrooms which allow for diffusion of oxygen and nutrients into imbedded cells. The gene expression shifts with the state in which microorganisms grow. As planktonic cells genes for motility structures are expressed while as biofilms, genes associate with glycolysis are expressed. Contrary to opinions held earlier that biofilms are microbial cells populated in an unorganised mesh of slime, biofilms are well controlled and organised mode cell growth.
As the biofilm matures, environmental conditions fluctuate some of which are beneficial for sessility and some which are not such that dispersal become inevitable. Biofilm dispersal can be caused by several factors such as nutrient deprivation, oxygen fluctuations and increase in toxic metabolites. There are several other stress-induced factors such as increasing concentrations of iron, carbon, nitrogen, Cyclic di-GMP and metabolites such as amino acids and activities of EPS- degrading enzymes which may favour dispersal of biofilms (Kostakioti, Hadjifrangiskou, and Hultgren 2013; Rowe, Withers, and Swift 2010)
Detachment of biofilms
Molecule studies have shown that bacteria depend on surfactant peptides secreted during quorum sensing to detach and disseminate(Otto 2013). Environmental factor such as temperature and nutrient concentration have their own influence on detachment of biofilms. In a study carried by (Morimatsu et al. 2012) high temperatures of 200 and 300C were observed to encouraged biofilm detachment which did not happen at lower temperatures of 50 and 100C. At low nutrient concentrations, biofilms detached after maturity regardless of temperature levels. The detachment of biofilms at
Biofilm formation is dependent on nutrient availability; thick biofilms are easily formed in nutrient-rich environments while bacteria hardly attach and form biofilms at very low nutrient concentrations(Costerton et al. 1995).
Role of exopolysaccharides on biofilm formation
Exopolysaccharides have impact on the structure of biofilms formed. Alginate, Pel and Psl have been shown to influence in biofilm formation pseudomonas spp (Ghafoor, Hay, and Rehm 2011). Alginates are linear polyanionic exopolysaccharides composed of uronic acids. Studies using mutants deficient in the production of one or more of these polysaccharides were generated showed that alginate-deficient mutants resulted in decreased number of viables cells while impacting the biofilm structure with intensified extracellular DNA (eDNA)-containing surface structures. The peculiar mushroom-like structure is determined by alginate in combination with Psl which determines the biofilm cell density. Mutants lacking in Psl and Pel are unable to form biofilms. Pel genes are the primary genes in the making of a glucose-rich matrix material required for the formation (Friedman and Kolter 2004) and in cell-cell signalling and enhanced resistance to aminoglycoside antibiotics (Colvin et al. 2011). Psl polysaccharides which is made up of mannose and galactose also mediate initial attachment and biofilm maturation (Ma et al. 2009; Ma et al. 2006).
Biofilm formation in Campylobacter jejuni is mediated by flagella-mediated adherence and its maturity by eDNA. Mutation in the genes encoding for these two results in strains with reduced tolerance to stress(Svensson, Pryjma, and Gaynor 2014)
Mixed biofilms
In nature microorganism exist in mixed cultures of which may consist of bacteria, fungi, protozoa algae and these organism impact on each by interactions which can prove difficult to determine in the laboratory. Within these large communities may exist a vast array of species and genera of microorganism most of which may be culturable and some viable but none culturable. This phenomenon brings to bear on the results obtained in the laboratory and the judgements we reach on how maximize the beneficial potentials of these organisms or as the case maybe in infection and contaminations eradication. With the discovery of biofilms and its existence in mono- and poly-colonies a lot remain to be understood.
Some proteins necessary for adhesion and biofilm formation in bacteria
In Legionella pneumophila (Mallegol et al. 2012) have demonstrated the importance of a glycosaminoglycan (GAG)-binding adhesin of L.pneumonia without which the organism cannot form biofilms. Mutants lacking in the gene encoding for this protein are unable to attach to surfaces and form biofilms which critical in its infection.
Quorum sensing is a cell density dependent inter cell communication medium via inducers to induce or repress the expression of genes involved in biofilm maturation and detachment.
Different models used in the study of biofilms
Overthe years since the discovery of biofilms scientists have come up with different in vitro and in vivo models for the study of biofilms. These models have often been designed to mimic typical biofilms either in the environment or in infections. The various models are discussed below:
Control of biofilms
There have been several suggestions to the control of biofilms in environment. Several surfaces developed have shown the ability of preventing biofilm formation. Copper and zinc oxide coating on surfaces demonstrate inhibitory properties on bacteria attachment. The applicability of these surfaces to all surfaces poses a problem as some surfaces may be required soft and some required hard.
Cleaning agents can be used in the effective control of microorganism attachment to surfaces which is largely dependent on the content of the cleaning agent. It is important factors which enhance bacteria adhesion in any way are eliminated or minimised in the making of these agent. For instance acidic ions have shown to increase the adhesive properties of microorganisms, it therefore become imperative that alkaline ions are favoured in the making of these agents.
In vitro biofilm models
In vivo biofilm models
My Null and alternative hypothesis
Environment has been shown a source of pathogen causing infections in the healthcare environments and one means of survival of pathogens in these environment is by growing as biofilms. Studies have shown that biofilms can be isolated from many surfaces in the hospital. The models used in the studies of these biofilms unfortunately differ so much from what is obtainable in nature. While the natural environment is characterised by dryness and limited nutrition due to the constant cleaning and drying in healthcare environments, models used provide moisture and nutrient rich environments.
My hypothesis is that growing dry biofilms in 24 well plates is reproducible, dry biofilms are different from wet biofilms in structure and in response to antimicrobial agents.
Detachment of biofilms
Molecule studies have shown that bacteria depend on surfactant peptides secreted during quorum sensing to detach and disseminate(Otto 2013). Environmental factor such as temperature and nutrient concentration have their own influence on detachment of biofilms. In a study carried by (Morimatsu et al. 2012) high temperatures of 200 and 300C were observed to encouraged biofilm detachment which did not happen at lower temperatures of 50 and 100C. At low nutrient concentrations, biofilms detached after maturity regardless of temperature levels. The detachment of biofilms at
Biofilm formation is dependent on nutrient availability; thick biofilms are easily formed in nutrient-rich environments while bacteria hardly attach and form biofilms at very low nutrient concentrations(Costerton et al. 1995).
Role of exopolysaccharides on biofilm formation
Exopolysaccharides have impact on the structure of biofilms formed. Alginate, Pel and Psl have been shown to influence in biofilm formation pseudomonas spp (Ghafoor, Hay, and Rehm 2011). Alginates are linear polyanionic exopolysaccharides composed of uronic acids. Studies using mutants deficient in the production of one or more of these polysaccharides were generated showed that alginate-deficient mutants resulted in decreased number of viables cells while impacting the biofilm structure with intensified extracellular DNA (eDNA)-containing surface structures. The peculiar mushroom-like structure is determined by alginate in combination with Psl which determines the biofilm cell density. Mutants lacking in Psl and Pel are unable to form biofilms. Pel genes are the primary genes in the making of a glucose-rich matrix material required for the formation (Friedman and Kolter 2004) and in cell-cell signalling and enhanced resistance to aminoglycoside antibiotics (Colvin et al. 2011). Psl polysaccharides which is made up of mannose and galactose also mediate initial attachment and biofilm maturation (Ma et al. 2009; Ma et al. 2006).
Biofilm formation in Campylobacter jejuni is mediated by flagella-mediated adherence and its maturity by eDNA. Mutation in the genes encoding for these two results in strains with reduced tolerance to stress(Svensson, Pryjma, and Gaynor 2014)
Staphylococcus aureus
S.aureus is a Gram positive non motile bacterium occurring in bunches that is often associated with wounds and surgery.
Mixed biofilms
In nature microorganism exist in mixed cultures of which may consist of bacteria, fungi, protozoa algae and these organism impact on each by interactions which can prove difficult to determine in the laboratory. Within these large communities may exist a vast array of species and genera of microorganism most of which may be culturable and some viable but none culturable. This phenomenon brings to bear on the results obtained in the laboratory and the judgements we reach on how maximize the beneficial potentials of these organisms or as the case maybe in infection and contaminations eradication. With the discovery of biofilms and its existence in mono- and poly-colonies a lot remain to be understood.
Some proteins necessary for adhesion and biofilm formation in bacteria
In Legionella pneumophila (Mallegol et al. 2012) have demonstrated the importance of a glycosaminoglycan (GAG)-binding adhesin of L.pneumonia without which the organism cannot form biofilms. Mutants lacking in the gene encoding for this protein are unable to attach to surfaces and form biofilms which critical in its infection.
Quorum sensing is a cell density dependent inter cell communication medium via inducers to induce or repress the expression of genes involved in biofilm maturation and detachment .
Different models used in the study of biofilms
Overthe years since the discovery of biofilms scientists have come up with different in vitro and in vivo models for the study of biofilms. These models have often been designed to mimic typical biofilms either in the environment or in infections. The various models are discussed below:
In vitro biofilm models
In vivo biofilm models
My Null and alternative hypothesis
Environment has been shown a source of pathogen causing infections in the healthcare environments and one means of survival of pathogens in these environment is by growing as biofilms. Studies have shown that biofilms can be isolated from many surfaces in the hospital. The models used in the studies of these biofilms unfortunately differ so much from what is obtainable in nature. While the natural environment is characterised by dryness and limited nutrition due to the constant cleaning and drying in healthcare environments, models used provide moisture and nutrient rich environments.
My hypothesis is that growing dry biofilms in 24 well plates is reproducible, dry biofilms are different from wet biofilms in structure and in response to antimicrobial agents.
Materials and methods
Standardization of cells
From a single colony of S.aureus on a nutrient agar plate an overnight culture was made in 10 ml of Luria broth. 1ml of the overnight broth was taken into eppendoff tube and centrifuged at 1000 rpm for 5 min and spent culture was aspirated. Cells were reconstituted in 1ml of phosphate buffer saline (PBS) vortexed and centrifuged and the procedure was repeated three times. Washed cell were adjusted to optical densities (OD) of 0.5. 0.4 0.3 0.2 and 0.1 at 570nm in separated eppendoff tubes. After appropriate serial dilution 20µl of diluents of each OD value was plated out by Miles and Misra on Nutrient agar plates and incubated overnight at 37 0C. The counts obtained were used in plotting a standard graph of OD against log values of cell.
Dry biofilm model.
A modification of the CDC biofilm model(Almatroudi et al. 2015) was used and used to grow dry biofilms. Iml of cells adjusted to 1×108cells/ml in 5% of TSB were inoculated into 24 well Corning™ Costar™ Flat Bottom Cell Culture Plates. Two sets of this model were made and a set incubated at room temperature and another at 250C for 288h (12days) with alternating cycles of dehydration and hydration at different time points as was in the CDC model. After 48 h of incubation broth was removed gently from the wells and allowed to dry for 48h at the temperature of incubation. at 250C. At the end of 48h of drying dry biofilms were hydrated with5 % TSB for 6h and the removed again for dehydration for 66h at the appropriate temperature. This is followed by another cycle of hydration for 6h, 42h of dehydration, 6h of hydration and 66h of dehydration. At the time point 96, 168, 216 and 288h, CFU, viability, biomass, biofilm structure, protein and polysaccharide content were analysed.
Quantification of biofilms
a. Cell count (CFU): Biofilms are washed three times with PBS. I ml of 1M NaCl is added to the well containing the biofilm and allowed to soak for 30min after which biofilms are scrapped carefully with the tip of I ml pipette to remove all biofilms. Using same pipette, the scrapings are removed into an Eppendorf tube. The tube is then sonicated for 5min in an ultrasonic bath for 5min, vigorously shaken vortexed for 2 min, followed by a sequential tenfold dilution in PBS and plating out on NA.
The remaining content in the Eppendorf tube is centrifuged for 10min at 6000rpm/min. the supernatant is removed and filtered through a syringe filter. The filtrate is used for protein and polysaccharide quantification.
b. Biomass. 1 ml of 0.1% crystal violet is added into washed biofilms in well and incubated at room temperature for 30min. the dye is discarded and the well washed carefully with distilled water and drained. 750 µl of absolute ethanol is added and incubated for 15 min. 100µl of the ethanol is removed into clean 96 well plates after carefully mixing by agitation and absorbance read at 570nm.
a. Bradford assay: The protein component of the recovered ECM was estimated by adding 0.5 ml to 0.5 ml of Bradford reagent in Eppendorf tubes and allowed to react for 10 minutes. The absorbance of 100 µL of the reaction mixture was measure at 595 nm in flat bottom 96-well plate using a Qiagen 96-well plate reader. The concentration of the protein component in the recovered ECM was estimated using a BSA standard graph.
b. Phenol-sulphuric assay: phenol sulphuric assay according to (Masuko et al. 2005) was used to quantify the polysaccharide component of biofilm ECM. 150 µL of concentrated sulfuric acid was added to 50 µL of recovered ECM sample in a 96-well microplate. 30 µL of 5% phenol was added and the mixtures were mixed and incubated for 5 min at 900C in a static water bath. The plates were thereafter allowed to cool to room temperature, wiped dry and the absorbance estimated at A492 nm. The polysaccharide component was estimated using a glucose standard graph.
Measuring of proliferation of S. aureus using alamarBlue® by spectrophotometry
References
Abelson, MB, and J McLaughlin. 2012. ‘Of biomes, biofilm and the ocular surface’, Rev Ophthalmol, 19: 52-4.
Almatroudi, Ahmad, Honghua Hu, Anand Deva, Iain B Gosbell, Anita Jacombs, Slade O Jensen, Greg Whiteley, Trevor Glasbey, and Karen Vickery. 2015. ‘A new dry-surface biofilm model: an essential tool for efficacy testing of hospital surface decontamination procedures’, Journal of microbiological methods, 117: 171-76.
Bales, Patrick M, Emilija Miljkovic Renke, Sarah L May, Yang Shen, and Daniel C Nelson. 2013. ‘Purification and characterization of biofilm-associated EPS exopolysaccharides from ESKAPE organisms and other pathogens’, PLoS One, 8: e67950.
Chandki, Rita, Priyank Banthia, and Ruchi Banthia. 2011. ‘Biofilms: A microbial home’, Journal of Indian Society of Periodontology, 15: 111.
Colvin, Kelly M, Vernita D Gordon, Keiji Murakami, Bradley R Borlee, Daniel J Wozniak, Gerard CL Wong, and Matthew R Parsek. 2011. ‘The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa’, PLoS Pathog, 7: e1001264.
Costerton, J William, Zbigniew Lewandowski, Douglas E Caldwell, Darren R Korber, and Hilary M Lappin-Scott. 1995. ‘Microbial biofilms’, Annual Reviews in Microbiology, 49: 711-45.
Donlan, Rodney M. 2001. ‘Biofilm formation: a clinically relevant microbiological process’, Clinical Infectious Diseases, 33: 1387-92.
———. 2002. ‘Biofilms: microbial life on surfaces’, Emerg Infect Dis, 8.
Fanning, Saranna, and Aaron P Mitchell. 2012. ‘Fungal biofilms’, PLoS Pathog, 8: e1002585.
Friedman, Lisa, and Roberto Kolter. 2004. ‘Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms’, Molecular microbiology, 51: 675-90.
Garrett, Trevor Roger, Manmohan Bhakoo, and Zhibing Zhang. 2008. ‘Bacterial adhesion and biofilms on surfaces’, Progress in Natural Science, 18: 1049-56.
Ghafoor, Aamir, Iain D Hay, and Bernd HA Rehm. 2011. ‘Role of exopolysaccharides in Pseudomonas aeruginosa biofilm formation and architecture’, Applied and environmental microbiology, 77: 5238-46.
Henrici, Arthur T. 1933. ‘Studies of freshwater bacteria: I. A direct microscopic technique’, Journal of bacteriology, 25: 277.
Heukelekian, H, and A Heller. 1940. ‘Relation between food concentration and surface for bacterial growth’, Journal of bacteriology, 40: 547.
Høiby, Niels. 2014. ‘A personal history of research on microbial biofilms and biofilm infections’, Pathogens and disease, 70: 205-11.
Janissen, R, DM Murillo, B Niza, PK Sahoo, MM Nobrega, CL Cesar, ML Temperini, HF Carvalho, AA de Souza, and MA Cotta. 2015. “Spatiotemporal distribution of different extracellular polymeric substances and filamentation mediate Xylella fastidiosa adhesion and biofilm formation. Sci Rep 5: 9856.” In.
Kafil, Hossein Samadi, and Ashraf Mohabati Mobarez. 2015. ‘Assessment of biofilm formation by enterococci isolates from urinary tract infections with different virulence profiles’, Journal of King Saud University-Science, 27: 312-17.
Kokare, CR, S Chakraborty, AN Khopade, and KR Mahadik. 2009. ‘Biofilm: Importance and applications’, Indian Journal of Biotechnology, 8: 159-68.
Kostakioti, Maria, Maria Hadjifrangiskou, and Scott J Hultgren. 2013. ‘Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era’, Cold Spring Harbor perspectives in medicine, 3: a010306.
Ma, Luyan, Matthew Conover, Haiping Lu, Matthew R Parsek, Kenneth Bayles, and Daniel J Wozniak. 2009. ‘Assembly and development of the Pseudomonas aeruginosa biofilm matrix’, PLoS Pathog, 5: e1000354.
Ma, Luyan, Kara D Jackson, Rebecca M Landry, Matthew R Parsek, and Daniel J Wozniak. 2006. ‘Analysis of Pseudomonas aeruginosa conditional psl variants reveals roles for the psl polysaccharide in adhesion and maintaining biofilm structure postattachment’, Journal of bacteriology, 188: 8213-21.
Mack, WN, JP Mack, and AO Ackerson. 1975. ‘Microbial film development in a trickling filter’, Microbial ecology, 2: 215-26.
Mallegol, Julia, Carla Duncan, Akriti Prashar, Jannice So, Donald E Low, Mauricio Terebeznik, and Cyril Guyard. 2012. ‘Essential roles and regulation of the Legionella pneumophila collagen-like adhesin during biofilm formation’, PLoS One, 7: e46462.
Martinez, Juan J, Matthew A Mulvey, Joel D Schilling, Jerome S Pinkner, and Scott J Hultgren. 2000. ‘Type 1 pilus‐mediated bacterial invasion of bladder epithelial cells’, The EMBO journal, 19: 2803-12.
Masuko, Tatsuya, Akio Minami, Norimasa Iwasaki, Tokifumi Majima, Shin-Ichiro Nishimura, and Yuan C. Lee. 2005. ‘Carbohydrate analysis by a phenol–sulfuric acid method in microplate format’, Analytical biochemistry, 339: 69-72.
Morimatsu, Kazuya, Kodai Eguchi, Daisuke Hamanaka, Fumihiko Tanaka, and Toshitaka Uchino. 2012. ‘Effects of temperature and nutrient conditions on biofilm formation of Pseudomonas putida’, Food Science and Technology Research, 18: 879-83.
Nadell, Carey D, Knut Drescher, Ned S Wingreen, and Bonnie L Bassler. 2015. ‘Extracellular matrix structure governs invasion resistance in bacterial biofilms’, The ISME journal, 9: 1700-09.
O’toole, George A, and Roberto Kolter. 1998. ‘Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development’, Molecular microbiology, 30: 295-304.
Otto, Michael. 2013. ‘Staphylococcal infections: mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity’, Annual review of medicine, 64: 175-88.
Rabin, Nira, Yue Zheng, Clement Opoku-Temeng, Yixuan Du, Eric Bonsu, and Herman O Sintim. 2015. ‘Biofilm formation mechanisms and targets for developing antibiofilm agents’, Future medicinal chemistry, 7: 493-512.
Rice, Louis B. 2008. ‘Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE’, Journal of Infectious Diseases, 197: 1079-81.
Rowe, Maria C, Helen L Withers, and Simon Swift. 2010. ‘Uropathogenic Escherichia coli forms biofilm aggregates under iron restriction that disperse upon the supply of iron’, FEMS microbiology letters, 307: 102-09.
Sheng, Guo-Ping, Han-Qing Yu, and Xiao-Yan Li. 2010. ‘Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: a review’, Biotechnology advances, 28: 882-94.
Svensson, Sarah L., Mark Pryjma, and Erin C. Gaynor. 2014. ‘Flagella-mediated adhesion and extracellular DNA release contribute to biofilm formation and stress tolerance of Campylobacter jejuni’, PLoS ONE, 9: e106063.
Vasudevan, Ranganathan. 2014. ‘Biofilms: microbial cities of scientific significance’, J Microbiol Exp, 1: 00014.
Watnick, Paula, and Roberto Kolter. 2000. ‘Biofilm, city of microbes’, Journal of bacteriology, 182: 2675-79.
Zhurina, MV, AV Gannesen, EL Zdorovenko, and VK Plakunov. 2014. ‘Composition and functions of the extracellular polymer matrix of bacterial biofilms’, Microbiology, 83: 713-22.
Zobell, Claude E. 1943. ‘The effect of solid surfaces upon bacterial activity’, Journal of bacteriology, 46: 39.
Otter, J. (2015).https://reflectionsipc.com/2015/06/30/biofilms-make-the-hospital-environment-far-from-inanimate/ Date access 11/09/2017
http://www.contagionlive.com/news/researchers-study-mdro-presence-in-biofilms-collected-from-hospitals-in-4-countries?sthash.6i4Bsc5k.mjjo/ Date access 11/09/2017
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


DMCA / Removal Request
If you are the original writer of this assignment and no longer wish to have your work published on UKEssays.com then please click the following link to email our support team:
Request essay removal